Джошуа Ледерберг в 1952 году ввел понятие «плазмида». Он обнаружил в кишечной палочке, кроме основной спиралевидной, вытянутой во весь свой немалый рост ДНК, еще и маленькие, свернутые в кольцо ДНК. О плазмидах заговорили медики, когда в 1959 году было доказано, что неэффективность многих антибиотиков обусловлена плазмидами, несущими гены устойчивости к антибиотикам. Плазмиды легко переходят от бактерии к бактерии, делая их невосприимчивыми к лекарствам. К примеру, вырабатываемый плазмидами фермент пенициллаза разрушает пенициллин, спасая бактерии от гибели. Что, конечно же, осложняет лечение больных.
Детальные наблюдения характера возникновения опосредованной плазмидами устойчивости сделаны в Японии и Англии. В 1945 г. в Японии для борьбы с дифтерией стали применять сульфаниламид. Он был высокоэффективен только первые 5 лет. Вскоре появились устойчивые штаммы дифтерийной палочки, а уже некоторое время спустя 80-90% изолятов были устойчивыми. Затем сульфаниламид заменили антибиотиками. Но уже в 1952 г. от больного дифтерией был выделен штамм дифтерийной палочки, одновременно устойчивый к тетрациклину, стрептомицину и сульфаниламиду. А в 1964 г. половина всех бактериальных штаммов, выделенных из больных дифтерией, несла гены устойчивости одновременно к четырем антибиотикам. Эти гены устойчивости были собраны в одной плазмиде, способной распространяться среди бактерий горизонтально.
Гены плазмид, в свою очередь, могут перекочевывать на хромосомы клеток-реципиентов. Считают, что таким путем в кишечную палочку попали гены, кодирующие ферменты инактивации антибиотиков. Возможно, эти гены впервые появились у почвенных бактерий, живущих рядом с грибами-продуцентами антибиотиков. В почвенных бактериях в геноме плазмид находятся детерминанты устойчивости к тяжелым металлам. Из природных резерватов плазмиды с транспозонами, несущие гены устойчивости, попадают к бактериям животных и человека и с помощью их распространяются по всему миру.
То, что затрудняло работу медиков, пригодилось генным инженерам. А им нужны были переносчики реконструированных молекул ДНК в живые объекты. Правда, вначале на эту роль прочили вирусы-бактериофаги. Но, проникнув в клетку, вирус ведет себя как опасный хищник. Он переключает ресурсы клетки на удовлетворение своих нужд и примерно через полчаса губит ее. Иначе поступает плазмида — в отличие от вируса, она не убивает клетку-хозяйку. Плазмида и приютившая ее клетка осуществляют симбиоз. Плазмида защищает бактерию от, например, пенициллина. Клетка предоставляет плазмиде ресурсы для питания, размножения. Все эти особенности симбиоза бактерий и плазмид (особенно способность плазмид переходить «из рук в руки», от одной клетки к другой) оказались источниками генно-инженерных методов.
Плазмидами Д. Ледерберг предложил обозначать все внеядерные генетические элементы, способные к автономной репликации. Сюда входят «каппа-частицы» у парамеций, экзогенные вирусы и т.д. Взгляд на плазмиды как на симбионты и альтернативный взгляд на них как на составную часть генома, согласно Д. Ледербергу, зависит от того, насколько широко исследователь трактует границы генома и наследственной системы организма.
Именно особенности жизненных циклов вирусов, плазмид и бактерий привело в дальнейшем к созданию так называемых «векторов» — искусственно сконструированных молекул наследственности, которые могут переносить чужеродный материал от одного организма к другому.
Какие генно-инженерные приемы подсмотрены в природе
Благодаря перемещающимся элементам, генофонды всех организмов потенциально составляют общий генофонд всего живого. Реализация этой потенции, т.е. передача генов между разными таксонами, детально документирована. У млекопитающих и птиц практически идентичные провирусные последовательности появились уже после их эволюционного обособления. Поток генов между далекими организмами является реальностью. Вопрос заключается только в том, насколько часто организмы присваивали чужеродные гены в качестве «благоприобретенных собственных функциональных генов».
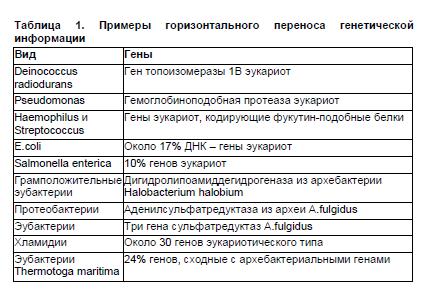
Наглядным примером горизонтального обмена между прокариотами и высшими эукариотами являются результаты исследований представленности в прокариотических геномах нуклеотидных последовательностей, типичных для эукариот (табл.1). Из этой таблицы следует, в частности, что в геноме симбионта человека, кишечной палочки, примерно 17% ДНК имеет эукариотическое происхождение. Хорошим примером естественной генетической трансформации является агробактериальная трансформация растительных клеток. Отличительная черта бактерий рода Agrobacterium (A.tumefaciens, A.rhizogenes) — способность вызывать развитие так называемых корончатых галлов (своего рода опухолей) у большого круга двудольных растений. При этом происходит перенос фрагмента ДНК агробактерии в геном растительных клеток. Такая клетка со встроенным участком агробактериальной ДНК продуцирует ряд органических веществ, служащих для агробактерии специфическими источниками углерода и азота. Такой перенос — уникальный природный процесс обмена генетической информацией между бактерией и растением, и именно его ученые взяли на вооружение для получения трансгенных растений, встраивая целевой ген в участок агробактериальной ДНК, переносимый в растение.
Эффективность трансформации растительных клеток может быть увеличена за счет использования штаммов A.tumefaciens, обладающих повышенной вирулентностью по отношению к данному виду растений. Тем не менее, частота трансформации весьма низка — только одна из 10 тысяч растительных клеток становится носителем рекомбинантной ДНК.
Возможно около 20 способов проникновения и межвидовой миграции генетических элементов, в их числе трансформация, трансдукция, перемещение транспозонов, плазмид, вирусов, неполовой обмен хромосомами и образование симбиотических ассоциаций. Информационная емкость переноса информации, выраженная в генах, варьирует от единиц до сотен и тысяч в случае плазмид и симбионтов.
Под влиянием стресса резко увеличиваются частоты горизонтального обмена материалом наследственности между бактериями, у растений — частоты перекрестного опыления у самоопылителей. В последние годы в различных моделях стресса у многих видов высших организмов наблюдают увеличение частот рекомбинационных событий, транспозиций, различных мутационных событий. Отчетливые данные о связи дестабилизации генетического материала с действием стрессирующих факторов, полученнные Б. МакКлинток, впоследствии привели к развитию представлений о системах «природной генетической инженерии» (Shapiro. 1992,1995).
Изначально термин «генетическая инженерия» применяли для обозначения целенаправленной манипуляции наследственными детерминантами с целью изменения существующих видов. В настоящее время этим термином обычно обозначают генетические манипуляции, с помощью которых формируется организм, имеющий новую комбинацию наследуемых признаков. Иначе ДНК-технологии можно определить как отрасль биологии, которая изучает явления и конструирование наследственности и изменчивости. Современный этап ДНК-технологий неразрывно связан с необходимостью увеличения источников благосостояния и здоровья человечества. ДНК-технологии стремительно увеличивают наши знания в одной из наименее исследованных областей — наследственности и законов ее изменения естественным и экспериментальным путем.
Свыкшись с материальностью генов, человек, естественно, тут же захотел заняться генной хирургией. Для этого в природе имеются ферменты рестриктазы, с высокой точностью разрезающие молекулу ДНК в определенных сайтах (сочетаниях нуклеотидов), и ферменты лигазы, «сшивающие» такие разрывы. Именно эти ферменты послужили основой для создания строго запланированных генных конструкций.
Использование рекомбинантных (перестроенных) ДНК различного происхождения составляет основу ДНК-технологий. Теоретически все 30-40 тысяч структурных генов человека и животных доступны теперь экспериментальному анализу. Поэтому желательна идентификация всех генов; составление карты тканеспецифичности их экспрессии; идентификация регуляторных областей генов; построение глобальной регуляторной карты генома; классификации генов по структурным и биохимическим функциям их продуктов; идентификация всех потенциальных белков и доменов; анализ распределения полиморфизма и мутаций; определение эволюционных и популяционных взаимосвязей; создание коллекции генетического материала и тд.