Именно природа этой дивергенции и есть "атомное ядро", тайну которого надлежит открыть. Для начала анализа ясно лишь, что, будучи процессом биологическим, она в то же время имела нечто отличающее ее от всякой другой дивергенции в живой природе. К тому немногому, что мы достоверно об этом знаем, принадлежит необычная быстрота данного ароморфоза – отпочкования нового, прогрессивного вида. Отсюда можно сделать вывод, что между обоими дивергирующими видами должны были существовать и крайне напряженные экологические отношения. Этого не было бы, если бы дивергенция с самого начала сопровождалась размежеванием ареалов. Вероятнее, напротив, что в пределах общего ареала происходило крутое размежевание экологических ниш.
Но главный вывод, который мы должны извлечь из стремительности дивергенции (а ее отрицают только немногие антропологи вроде А. Валлуа, держащиеся слабо обоснованной концепции происхождения Homo sapiens не из того или иного вида палеоантропов, а из "пресапиенсов", восходящих к среднему или даже раннему плейстоцену), состоит в том, что перед нами продукт действия какого-то особого механизма отбора.
Прежде всего хотелось бы реконструировать не только само раздвоение или, вернее, отпочкование, но и его более мелкие промежуточные уровни. Мыслимо ли это?
Но следует ли вообще думать, что палеонтология всегда ищет и находит все промежуточные ступеньки между одной биологической формой и филогенетически последующей, уже существенно отличающейся? Состоит ли сама идея палеонтологии в том, что в принципе должны где-то существовать останки всех мыслимых степеней сочетания прежнего и нового? Нет, конечно, в этом филогенетическом переходном мосту всегда много неустойчивых, хрупких образований, не надстраивающихся в чисто количественном ряду друг над другом, а представляющих очень бедные по числу, очень вариативные и очень ломкие образования. Пока, наконец, одно из них не станет основанием для жизнеспособной, многочисленной ветви.
Палеонтологи иногда называют это практически неведомое им, исчезнувшее соединение эволюционных форм "черешком". Этот черешок, на котором держится новый вид, всегда тонок, почти никогда не доступен прямому изучению палеонтологии. Иначе говоря, в диапазоне между родительскими и нашедшими свою почву стойкими, дающими богатые соцветия таксономическими единицами находится обвал возникавших и гибнувших нежизнеустойчивых форм. В десятки раз труднее изучить этот "черешок" ответвления человека – Homo sapiens, оторвавшегося относительно быстро на огромную, как мы уже знаем, биологическую дистанцию: на расстояние нового семейства. Уж очень специфично то, что возникло: вид, отличающийся инверсией процессов высшей нервной деятельности, – "животное наоборот". Посмотрим, что же мы все-таки имеем в руках из костного материала, годного для непосредственной датировки и биологической фиксации дивергенции.
В результате блестящих исследований ископаемых эндокранов, осуществленных В. И. Кочетковой, мы узнали нечто более важное, чем существование тут и там в четвертичных отложениях "переходных" черепов, расположенных по сумме признаков на том или ином отрезке пути между "неандертальцем" и "кроманьонцем". Открытие Кочетковой состоит в глубоком изменении прежнего представления о самих кроманьонцах, т.е. об ископаемых неоантропах начальной поры верхнего палеолита, которые оказались не тождественными позднейшим неоантропам. Трудно переоценить огромность этого, казалось бы, тончайшего сдвига: кроманьонцы – не то, что привычно и долго о них воображали. А именно было общепринято, что кроманьонцы – это другое наименование для нас самих. Посади с нами за обеденный стол неандертальца – все согласны, что его общество было бы невыносимо; но посади кроманьонца (хорошо одетого, побритого, обученного нашему языку и манерам) – его якобы никто бы и не отличил. Соответственно подчас говорят: "Мы, кроманьонцы". Исследование эндокранов обнаружило тут ошибку. Трудно сказать, оценила ли в полной мере сама В. И. Кочеткова всю капитальность своего вывода, что черепа группы ископаемых неоантропов (Homo sapiens fossilis) серьезно отличаются – по крайней мере в некотором проценте экземпляров и тем самым в среднем – от величин типичных и устойчивых для ныне живущих неоантропов, т.е. людей современного типа. Мало того, выяснилось, что это отклонение характеризует людей первой половины верхнего палеолита (столь же неточно в общежитии именуемой "ориньяком"). Такие верхнепалеолитические индивиды, как Кро-Маньон III, Маркина Гора, оказались по эндокрану, т.е. по макроморфологии головного мозга, вообще ближе к палеоантропам, чем к неоантропам.
В своих цифровых таблицах различных параметров строения мозга В. И. Кочеткова убедительно выделила ископаемых неоантропов в особую группу, оказавшуюся глубоко специфичным перевалом в антропогенезе. Некоторые показатели, нарастающие во всей цепи от шимпанзе к австралопитекам и далее, достигают своей кульминации именно в группе ископаемых неоантропов, после чего кривая падает. Другие показатели, наоборот, достигают кульминации накануне появления этой группы, т.е. у палеоантропов, а с ископаемых неоантропов уже начинается нисходящая линия, характерная для неоантропов вообще по сравнению с ростом соответствующей кривой у троглодитид вообще. Однако следует помнить, что вся группа ископаемых неоантропов пока представлена сравнительно немногочисленными находками. Тем выразительнее выступает ее полиморфность (см. составленную мною сводную таблицу по опубликованным данным Кочетковой).
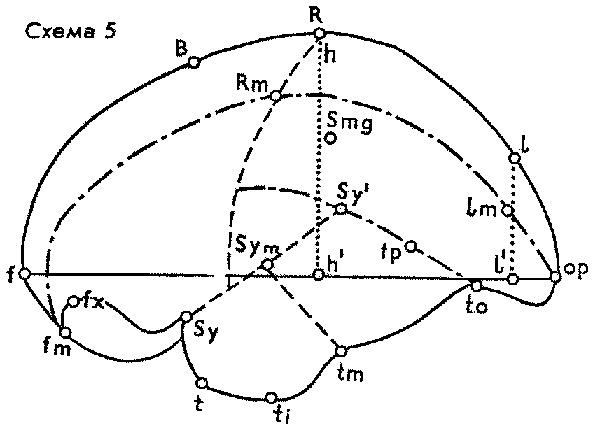
| Понгиды | Троглодитиды | Гоминиды | ||||||
| Шимпанзе | Австра | Археоантропы | Палеоантропы | Ископаемые неоантропы | Современные люди | |||
| лопитек | ||||||||
| Тотальный продольный размер | 106 (104-109) | 125 | 153,7 (145-159) | 174,7 (163-182) | 186.3 (176-196) | 167,1 (150-183) | ||
| Лобная доля | Верхний продольный диаметр fm – R | 81,3 (78-86) | 92,0 | 110,0 (98-118) | 128,7 (123-139) | 135,3 (98-147) | 129,4 (116-143) | |
| Нижний продольный диаметр fm – Sy | 33,6 (31-35) | 55,5 (51-60) | 61,4 (58-65) | 55,0 (46-60) | 49,0 (43-57) | |||
| Дуга нижнего лобного края fm – Sy | 56.3 (52-63) | 70,9 (61-79) | 83,8 (75-91) | 74,0 (71-76) | 70,2 (58-87) | |||
| Дуга латериального бугра fx–Sy | 28,0 (22-34) | 41.5 (35-50) | 52.9 (46-58) | 44,4 (39-50) | 43,8 (41-47) | |||
| Максимальный широтный размер р–р | 69,3 (66-75) | 89,0 (81-93) | 108,1 (103-115) | 108,0 (102-113) | 104,4 (89-126) | |||
| Теменная доля | Верхняя | Продольный диаметр R-1 | 31.3 (26-36) | 35,0 | 48,0 (48-51) | 57,5 (52-64) | 67,9 (60-76) | 65,8 (51-77) |
| Высотный диаметр калотты h–h' | 48,0 (44-51) | 54,7 | 66,0 (61-69) | 77,7 (72-82) | 76,6 (68-90) | 85,9 (72-97) | ||
| Нижняя | Rт–1т | 40,3 (33-57) | 49,0 | 67,7 (65-70) | 82,3 (74-91) | 95,1 (87-101) | 88,1 (79-101) | |
| Поперечный диаметр smg–smg | 71 (61-80) | 103,3 (94-112) | 118.9 (104-132) | 116.5 (101-140) | 124,1 (108-152) | |||
| Височная доля | Продольный диаметр t-tp | 62,6 (61-65) | 66,0 | 73,5 (73-74) | 86,6 (78-94) | 92,5 (90-95) | 90,4 (76-100) | |
| Поперечный диаметр eu–еu | 88,0 (86-89) | 123,3 (118-129) | 136,3 (116-146) | 136,3 (126-146) | 132.2 (117-158) | |||
| Высотный диаметр Sy–ti | 23,3 (21-25) | 34,0 (28-40) | 39,2 (34-46) | 47,0 (43-51) | 42,7 (33-51) | |||
| Высотный диаметр Sy'-to | 32,6 (31-34) 2 | 33,5 (33-34) | 42.2 (36-46) | 51,0 (46-5S) | 46.2 (38-54) | |||
| Высотный диаметр tm–Sym | 3,3 (21-23) | 32,0 (29-35) | 36,2 (32-41) | 37,4 (33-42) | 42,0 (36-51) | |||
| Затылочная доля | Продольный диаметр 1m–om | 38,6 (35-41) | 38,0 | 42,3 (38-47) | 44,9 (39-50) | 42,9 (35-55) | 38,0 (29-52) | |
| Высотный диаметр 1-1' | 25,0 (22-27) | 37,0 | 44.0 (38-50) | 50,9 (44-59) | 50,6 (37-56) | 47,9 (39-54) | ||
| Степень нависания затылка над мозжечком (в относительных величинах) | 3,5 (2,9-3,8) | 6.8 (6.3-7.3) | 10,9 (9.3-13,5) | 8,9 (5,6-13,5) | 5,4 (2,9-9,5) | |||
| То же (в баллах) | 0,7 (0.6-0,8) | 1.4 (1,3-1,5) | 2,1 (1,6-2.7) | 1,8 (1,2-2,7) | 1,1 (0,6-1.9) |