НЕЙРОФИЗИОЛОГИЯ ВКУСА
Функциональная роль вкусовой системы у позвоночных животных разных видов неодинакова. У наземных животных вкусовая рецепция связана с оценкой качества пищи или отвергаемых веществ, тогда как у рыб вкус играет роль дистантной рецепции и наряду с обонянием обеспечивает пространственную ориентацию. Это связано с тем, что вкусовые почки у рыб обнаруживаются за пределами ротовой полости и находятся на губах, жабрах, плавниках, голове, туловище. Вкусовая чувствительность играет большую роль в формировании поведения наземных млекопитающих. Благодаря этой чувствительности животные выбирают адекватные пищевые вещества. Однако обычно ощущение вкуса сопровождается тактильными и температурными характеристиками пищи, находящейся в ротовой полости, а также ее запахом. Сочетание всех этих раздражений создает ощущение вкуса и адекватную реакцию животного на пищевое вещество.
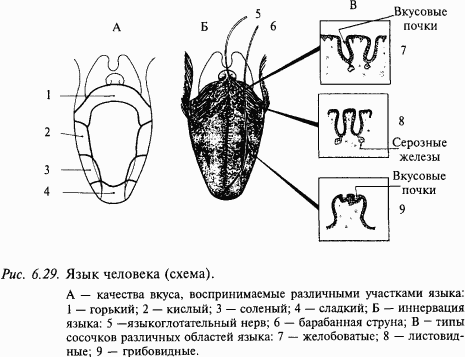
Вкусовая рецепция у человека осуществляется вкусовыми луковицами языка, которые встроены в более крупные структуры – сосочки. В каждом сосочке находится по нескольку вкусовых луковиц. Имеются следующие разновидности вкусовых сосочков: на кончике и по бокам языка – грибовидные, по бокам – листовидные, а у основания языка – желобоватые (рис. 6.29). Клетки вкусовых луковиц проходят перпендикулярно через всю толщу эпителия, достигая базальными концами мембраны, а в апикальной части они образуют вкусовой канал, соединенный с ротовой полостью через вкусовую пору. Вкусовая луковица состоит из 30–80 уплощенных, вытянутых веретенообразных клеток, тесно прилегающих друг к другу наподобие долек апельсина. Тонкие афферентные волокна (2/3 волокон не миелинизированы) образуют синаптические контакты на средней и базальной частях этих клеток. Если перерезать волокна, иннервирующие вкусовую луковицу, то последние полностью дегенерируют. Регенерация нерва ведет к восстановлению луковицы.
Пройдя через вкусовую пору, молекулы вещества попадают на верхушки вкусовых клеток, на апикальной поверхности которых имеются микроворсинки. В результате взаимодействия вкусовых молекул с мембраной клеток в последних возникает генераторный потенциал, а в соответствующих афферентных волокнах – импульсный разряд.
У млекопитающих, включая человека, первичный центр вкуса находится в ядре одиночного (солитарного) пути, который иннервируется ипсилатеральными волокнами лицевого, языкоглоточного и блуждающего нервов, по которым передается возбуждение от вкусовых рецепторов. Ядро одиночного пути находится в продолговатом мозге. Большинство волокон лицевого нерва оканчиваются в ростральной части, волокна языкоглоточного – в средней части, блуждающего – в средней и каудальной частях ядра. У млекопитающих ядро одиночного пути хорошо прослеживается на всем протяжении продолговатого мозга в дорсальной его части. Клеточное строение этого ядра напоминает строение желатинозной субстанции (Роланда) спинного мозга.
Вкусовые луковицы передних двух третей языка иннервируются язычным нервом (n. lingula) и барабанной струной (chorda tympani). Барабанная струна представляет собой аксоны клеток, тела которых находятся в коленчатом узле (ganglion geniculi). Центральный отросток этих клеток (нерв Врисберга) заканчивается в передней части ядра одиночного пути. По афферентным волокнам язычного нерва и барабанной струны проводится информация о вкусе, а также импульсация от температурных, тактильных и болевых рецепторов передней трети языка. От задней трети языка, твердого неба и миндалин информация о вкусе передается по волокнам языкоглоточного нерва. Кроме того, вкусовые луковицы глотки иннервируются волокнами блуждающего нерва (верхнегортанный нерв). Вкусовые афферентные волокна далее поступают в ствол мозга и заканчиваются на нейронах передней части ядра одиночного пути продолговатого мозга. Вкусочувствительная слизистая оболочка ротовой полости и глотки проецируется в ядро одиночного пути соматотопически.
От нейронов ядра одиночного пути одна часть волокон 2-го порядка направляется в составе медиального пучка к дугообразному ядру таламуса, а другая – к нейронам дорсальной части моста (парабрахиальное ядро). Аксоны таламических нейронов образуют корковую проекцию вкусового анализатора. Точная локализация коркового центра вкуса у человека не известна, хотя предполагают, что он тесно прилегает к корковой проекции ротовой полости. Считают, что таламокортикальная система обеспечивает вкусовое различение, тогда как связи с лимбическими структурами обеспечивают мотивационные характеристики пищевого поведения.
Электрофизиологические исследования последних лет показали, что, по-видимому, основная информация о вкусе передается по волокнам барабанной струны и языкоглоточного нерва. По данным этих исследований, практически все волокна барабанной струны и нейроны ядра одиночного пути продолговатого мозга реагируют повышением частоты импульсного разряда на несколько вкусовых модальностей, т.е. являются мультимодальными. Значительная часть нейронов продолговатого мозга реагирует не только на вкусовые модальности, но и на температурные и тактильные раздражители. Такую же широкую полимодальность обнаружили у таламических нейронов вкусового анализатора. Высказано преположение, что характеристика вкусовых раздражителей представлена относительной величиной активности многих нейронов. Каждый вкусовой раздражитель активирует нейроны соответствующей нейронной популяции в разной степени, причем «рельеф» этой активности характерен для каждого вкуса. Это своеобразный код вкусового качества: кодирование происходит как номером канала, так и относительной активностью в соответствующих каналах. Этот принцип, по-видимому, соблюдается для всех уровней переработки информации о вкусе.
Вопросы
1. Строение периферической части вкусового анализатора.
2. Пути и центры проведения информации о вкусе в головном мозге человека.
Литература
Основы сенсорной физиологии/Под ред. Р. Шмидта. М.: Мир, 1984
Глава 7
ВЫСШИЕ ФУНКЦИИ НЕРВНОЙ СИСТЕМЫ
Общепризнано, что нервная высшая деятельность человека и животных обеспечивается целым комплексом совместно работающих мозговых структур, каждая из которых вносит в этот процесс свой специфический вклад. Это означает, что нервная высшая деятельность в целом может нарушаться при поражении большого числа мозговых структур. Достаточно условно можно выделить три основных блока мозга, участие которых необходимо для осуществления любого вида психической деятельности [Лурия А. Р., 1973]:
• блок, обеспечивающий регуляцию тонуса головного мозга;
• блок получения, переработки и хранения информации;
• блок программирования, регуляции и контроля психической деятельности.
Каждый из этих блоков имеет иерархическое строение с верхним этажом в коре больших полушарий. Содержательное описание блоков в экспериментах на животных было сделано в соответствующих разделах учебника.
Блок тонуса головного мозга построен по принципу «неспецифической» нейронной сети, которая осуществляет свою функцию путем градуального изменения функционального состояния головного мозга и не имеет непосредственного отношения к приему и переработке поступающей информации. Одним из выдающихся достижений в этой области было установление того факта, что тонус коры больших полушарий мозга обеспечивается аппаратом не самой коры, а структурами мозгового ствола и подкорковыми отделами мозга. При этом подкорковые структуры не только тонизируют кору, но и сами испытывают ее регулирующее влияние. Эти аппараты головного мозга занимают в основном область мозгового ствола, промежуточного мозга и медиальную поверхность больших полушарий (подробнее см. выше).
Второй функциональный блок обеспечивает прием, переработку и хранение информации, поступающей в головной мозг по соответствующим сенсорным каналам. Нейронные сети этого блока работают не по принципу «градуальности», а осуществляют проведение и обработку строго специфичных сигналов. Эта специфичность начинается уже на периферии сенсорных систем: детекторами специфических энергий являются рецепторы. Другими словами, этот блок головного мозга обладает высокой модальной специфичностью. Его основу в коре больших полушарий составляют первичные, вторичные и третичные корковые зоны, расположенные в ретроцентральных участках коры больших полушарий (имеется в виду мозг приматов, включая человека). Первичные зоны этих областей коры характеризуются развитием IV слоя коры. Например, в первичном зрительном корковом поле VI (поле 17, по Бродману) IV слой дополнительно дифференцирован на три подслоя: IVa, IVb, IVc. Для вторичных и третичных корковых зон характерно то, что ведущее место занимают слои II и III. Второй блок имеет иерархическое строение, что соответствует и последовательности в переработке информации. Наиболее полно принцип иерархичности изучен на примере зрительного анализатора (подробнее см. главу 6, «Нейрофизиология зрительной системы»). Вместе с тем не вызывает сомнений, что принцип иерархичности нейроанатомической конструкции и последовательности обработки информации соблюдается также и в других анализаторах.