Восстановление показателей нейронной активности в период ПС имело частичный характер. Можно предполагать, что это восстановление отражает растормаживание нейронных механизмов, имеющих отношение к явлениям, специфическим для ПС, как, например, быстрым движениям глаз или возникновениям сновидений. Однако для такого разграничения имеющихся сведений недостаточно, их могут дать дальнейшие исследования.
На рис. 45 приведена сводная схема расположения в структурах мозга систем, участвующих в регуляции уровня бодрствования и глубины сна. В соответствии с этой схемой последовательное включение нейрогуморальных механизмов, обусловливающих засыпание, смены стадий сна и пробуждение, можно представить по сигналам наступления темноты, прекращения деятельности и обстановки приготовления ко сну.
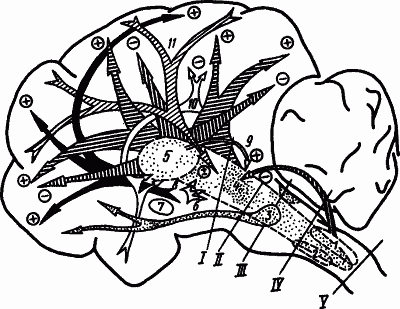
Рис. 45. Структуры мозга, участвующие в регуляции уровня бодрствования и глубины сна (по А.Н. Шеповальникову):
1 — синхронизирующая бульбарная система, 2 — «дополнительная» бульварная система, 3 — мостовой комплекс, обеспечивающий парадоксальный сон, 4 — активирующая ретикулярная формация ствола мозга, 5 — синхронизирующая таламическая система, 6 — активирующие влияния гипоталамуса на структуры ствола мозга, 7 — базальная синхронизирующая область, 8 — активирующие влияния гипоталамуса на структуры коры, 9 — облегчающее влияние высокочастотной стимуляции интраламинарных ядер таламуса на ретикулярную систему, 10 — влияния лимбической системы, способствующей сну, 11 — облегчающие и угнетающие влияния коры мозга на ретикулярную систему; I—V — уровни перерезок мозгового ствола при изучении нервных механизмов сна; знаком «+» обозначено активирующее действие, знаком «−» — угнетающее
Условное и безусловное торможение, которое развивается на фоне изменения активности гормонов и метаболизма, а также ограничение сенсорной информации и умеренное утомление обусловливают угнетение внешней деятельности путем ослабления влияния активирующих десинхронизирующих систем (рис. 45, 4, 6, 9, 11). Начинается засыпание.
Снижение тонуса активирующих систем помогает выявлению развивающейся к этому времени деятельности гипногенных синхронизирующих систем (рис. 45, 1, 2, 5, 7, 10). Гипоталамус оказывает трофотропное влияние на структуры ствола. В процесс вовлекается хвостатое ядро и утрачивается способность к активным движениям, блокируется поступление сенсорной информации. В нервных клетках ствола мозга, особенно в области синхронизирующей бульбарной системы, накапливается серотонин. Развивается медленноволновый сон.
Нарастающая на протяжении десятков минут медленноволнового сна деятельность гипногенных систем достигает степени, при которой (пока неясно, каким образом) происходит быстрый переход к активами центров дорсомедиальных отделов моста (рис. 45, 3), обусловливающих десинхронизацию в коре без участия ретикулярной формации среднего мозга. В соответствующих отделах мозга усиливается обмен веществ и повышается температура. Частичная активизация коры и ее связи с лимбической системой создают условия для появления сновидений. В структурах ствола мозга накапливается норадреналин и уменьшается серотонин. Возникает парадоксальный сон.
Активность нейрогуморальных механизмов парадоксального сна постепенно уменьшается и может на протяжении нескольких минут снизиться настолько, что усилившиеся за это время влияния гипногенных систем вызовут быструю обратную перестройку деятельности мозга в состояние медленноволнового сна. Однако это состояние, хотя и более продолжительное, чем состояние парадоксального сна, через некоторое время снова сменится им. Такое циклическое чередование стадий сна длится всю ночь, причем к утру продолжительность стадий парадоксального сна обычно возрастает.
С утренним рассветом связаны такие изменения активности гормонов и обмена веществ, которые в суточной периодике означают переход к активной внешней деятельности в светлое время. Под влиянием этих изменений, а также нарастающих потоков сенсорной информации и завершения восстановительных процессов в накануне работавших до утомления структурах мозга происходит снижение тонуса гипногенных систем, что благоприятствует резкому возрастанию деятельности активирующих систем, особенно десинхронизирующего действия ретикулярной формации среднего мозга. Наступает пробуждение.
В разные стадии сна по-разному протекают процессы высшей нервной деятельности. Так, выработанная на словесную инструкцию двигательная реакция сжатия руки в кулак сопровождалась изменениями ЭЭГ и ЭМГ, наиболее выраженными в ПС. Описаны нарушения памяти при устранении ПС. Однако приводятся доказательства того, что этим не исключается консолидация долгосрочной памяти при выработке реакций пассивного избегания.
В управлении сном принимает участие наряду с нервной гуморальная регуляция. Из аминокислотных остатков выделен дельта-пептид сна, вызывающий дельта-волны и характерный для МС (М. Монье, Г. Шененбергер, 1975). Из мозга животных, лишенных сна, выделен фактор S, увеличивающий продолжительность МС. В опытах на животных и в наблюдениях на людях показана роль серотонина и норадреналина в процессах сна. На основе этих данных предложен следующий механизм развития и прекращения сна: подавление симпатической активности приводит к поверхностному сну, расходование серотонина соответствует глубокому МС, расходование норадреналина — ПС, возбуждение симпатической активности вызывает пробуждение.
По мере накопления факторов и наблюдений за сном человека и животных возникали разные теоретические представления о его природе.
Теория гипнотоксинов. Всем известное освежающее действие сна наводило на мысль, что за это время организм освобождается от накопившихся при дневной деятельности токсических продуктов обмена веществ, которые и вызывают сонное торможение нервных клеток мозга. Доказательством такого представления о химической природе сна служили опыты, в которых сыворотка крови долго не спавшей собаки, введенная другой собаке, вызывала у нее сон (Р. Лежандр, А. Пьерон, 1910). Однако скоро выяснилось, что это неспецифическое действие белков любой сыворотки крови. Кроме того, теории гипнотоксинов противоречит тот факт, что сон можно превозмочь интересным занятием. Самое наглядное опровержение химической теории сна дало наблюдение за сросшимися близнецами: при общем кровообращении, т.е. в одной и той же химической среде одна голова может спать, в то время как другая — бодрствовать.
Вместе с тем недавно было показано участие гуморальных факторов в развитии сна. Из крови животного, заснувшего в результате раздражения гипногенных зон таламуса, был получен уже упоминавшийся выше пептид, введение которого вызывало сон. Такой пептид дельта-сна, извлеченный из экстракта ткани ствола мозга, оказался активнее, чем приготовленный из ткани коры.
Теория центров сна. Эта теория ведет начало от клинических наблюдений за больными энцефалитом, вызывающим летаргический сон. У этих больных оказывался воспаленным определенный участок ствола мозга, который стали рассматривать как центр сна (К. Экономо, 1930). Предположение, что сон вызывается возбуждением специальных центров мозгового ствола, получило поддержку в опытах с раздражением структуры промежуточного мозга, под действием которого кошка укладывалась в характерную позу сна и засыпала (В. Гесс, 1933). Однако дальнейшие исследования показали, что такой результат можно получить, раздражая в определенном режиме стимуляции различные структуры мозга, что противоречило представлению о нервном центре, который должен был иметь определенную локализацию. Кроме того, клинические наблюдения показали, что патология сна не связана с определенной локализацией поражения мозга. Вместе с тем вопрос о центрах сна представляет существенный интерес.