2. Перестраивается структура импульсных потоков. Непрерывно-ритмическое следование импульсов в состоянии бодрствования при переходе к МС сменяется их группированием в пачки, а при наступлении ПС следует частичное возвращение к непрерывно-аритмическому типу импульсации (рис. 44). Результаты анализа 900 отрезков записи фоновой активности корковых нейронов, каждого при состоянии бодрствования животного, во время МС и ПС представлены в табл. 8.
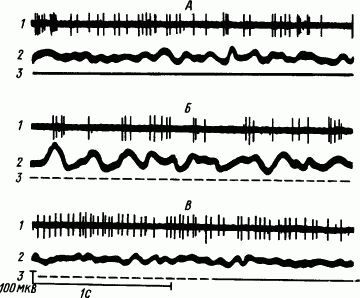
Рис. 44. Фоновая импульсация нейронов теменной коры мозга кошки в состоянии бодрствования (А), медленноволнового сна (Б) и парадоксального сна, прерываемого пробуждением (В):
1 — импульсная активность, 2 — электрокортикограмма, 3 — состояния животного (бодрствование — сплошная линия, сон — пунктир)
Таблица 8. Распределение различной структуры импульсных потоков теменной коры кошки в разные стадии сна, %

Во время МС каждая пачка объединяет 5–15 импульсов с интервалами молчания между пачками 200–500 мс. Такое формирование структуры импульсного потока характерно для влияний восходящей неспецифической таламокортикальной системы, приводящих к развитию тормозных процессов в коре. Во время ПС большинство нейронов возвращается к непрерывно-аритмическому типу фоновой импульсации. Однако часть нейронов сохраняет ее пачечно-групповую структуру, т.е. в этой стадии возникает некоторое разделение возбуждающих и тормозящих механизмов корковой деятельности.
3. Исследование взаимосвязи импульсации корковых нейронов ползало, что в медленноволновой стадии она резко усиливается, повышается синхронизация импульсных потоков нейронов, находящихся не только близко друг от друга, но и на таком расстоянии, при котором синхронизация их импульсации при бодрствовании была незначительной; в парадоксальной стадии она снова становится незначимой (табл. 9).
Таблица 9. Взаимосвязь импульсных потоков (по коэффициентам корреляции) разноудаленных нейронов теменной коры мозга кошки в разные стадии сна

Такая синхронность разрядов обширных популяций нейронов, видимо, также обусловлена диффузными влияниями восходящей таламокортикальной системы, организующей развитие процессов сонного торможения. Этим объясняется и то, что возникающие в МС пачки импульсов оказываются приуроченными к фазам дельта-волн. Аналогичные изменения корреляции импульсные разрядов корковых нейронов наблюдались при раздражении неспецифических структур таламуса, приводящем к возникновению веретен. При переходе к ПС таламические влияния ослабевают и усиливается активность структур среднего мозга.
4. При испытании реактивности нейронов теменной коры кошки оказалось, что в ответах на афферентное (слуховое, зрительное, тактильное) и в меньшей степени на прямое раздражение таламической радиации в медленноволновой и парадоксальной стадиях прогрессивно уменьшается число нейронов, реагирующих как возбуждением, так и торможением (табл. 10).
Таблица 10. Динамика реактивности корковых нейронов в процессе развития сна
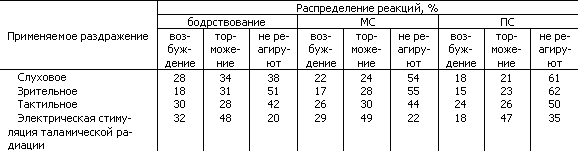
Существует некоторое различие в динамике ответов на афферентные раздражения и прямую стимуляцию входных в кору путей, в частности практически неизменный высокий процент заторможенных нейронов в одном случае и сравнительно малое число нереагирующих — в другом. Эти различия могут указывать на то, что развитие сна затрагивает и специфические релейные ядра таламуса, затрудняя прохождение через них афферентных сигналов. Нейроны, реагировавшие возбуждением при бодрствовании животного, могли при развитии сна не только перестать возбуждаться, но и начинали реагировать торможением. Примечательно, что в стадии МС уменьшается вариабельность импульсных реакций нейронов на раздражения, что можно объяснить как результат резкого ограничения полисенсорного реагирования.
5. Построение гистограмм распределения величин латентных периодов импульсных ответов раздельно для их значений менее и более 5 мс выявило, что прогрессирующее в МС и ПС уменьшение средней величины латентного периода импульсных ответов происходит не за счет ускорения проведения импульсов, а в результате уменьшения доли длиннолатентных реакций при неизменности коротколатентных. Если при бодрствовании последние составляли менее трети всех тветов, то в период ПС их было больше половины. Сон упрощает путь движения нервных процессов в коре.
6. Возникающий в ответ на раздражение сложный узор мозаики активности элементарных нейронных ансамблей с наступлением сна претерпевает глубокие изменения. В период МС функциональная мозаика элементарных нейронных ансамблей становится более грубой ввиду размывания границ между ними, приводящему к их слиянию, а в ПС происходит лишь некоторый сдвиг в сторону восстановления структуры мозаики.
Это наглядно показывает математическая реконструкция мозаик активности нейронов, построенная по данным одновременной мультиэлектродной регистрации нейронных реакций методом поэтапной оптимизации. При этом во время МС микроочаги возбуждающихся нейронов, ранее разделенные тормозящимися, сливаются между собой, образуя массивные зоны возбуждения наряду с массивами тормозящихся нейронов. В период ПС эти массивы дробятся, но не достигают степени дробности, характерной для состояния бодрствования.
7. Если считать, что нейроны передают информацию потоками импульсов, то можно попытаться оценить их информационную деятельность в понятиях теории информации, разработанной для технических систем связи (К. Шеннон, 1963). Результаты такой оценки представлены в табл. 11.
Таблица 11. Информационные характеристики импульсной активности нейронов теменной коры мозга кошки в разные фазы сна

В медленноволновую стадию по сравнению с состоянием бодрствования количество и особенно скорость передачи информации уменьшаются, хотя избыточность канала связи резко возрастает, а в парадоксальную стадию эти показатели вновь увеличиваются, превышая по пропускной способности наблюдавшиеся при бодрствовании. По-видимому, при засыпании выключаются некоторые источники потоков информации и нейронные каналы связи оказываются недозагруженными, что и обусловливает увеличение избыточности. Возрастание пропускной способности нервных каналов связи при переходе к ПС может отражать их мобилизацию для передачи информации, необходимой для деятельности, осуществляемой в эту стадию сна.
Все перечисленные особенности поведения корковых нейронов в разные стадии сна дают основание судить о нейрофизиологических механизмах его развития на нейронном уровне. Так, выключаемые из деятельности при засыпании нейроны, которые характеризуются полимодальностью, длинным латентным периодом и вариабельностью реакции, а также высокой частотой импульсации и повышенной возбудимостью, очевидно, имеют наиболее близкое отношение к осуществлению высших функций мозга. Можно предположить, что с их выключением связана потеря способности к активному восприятию окружающего и утрата сознания. Ограничение других функций мозга, возможно, обусловлено снижением возбудимости нейронов, замедляющих свои разряды, уменьшением информативности передаваемых сигналов, огрублением рабочих мозаик нейронной активности и перестройкой структуры импульсных потоков с преобладанием пачечного типа, характерного для восходящих таламокортикальных тормозных влияний.