При выработке условного рефлекса постановки лапы кошки на подставку исследовали реакции ЭЭГ и нейронной активности в моторной коре. Наблюдаемое при этом их единообразие в случае применения различных по характеру и интенсивности условных раздражителей привело к заключению, что сигнал лишь запускает программу ответного двигательного акта. Это происходит, когда наступает повышение эффективности синапсов на входах нейронов — генераторов команд.
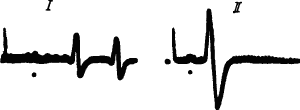
Рис. 23. Укорочение времени прохождения через метаторакальный ганглий таракана нервных импульсов, приводящих в движение его задние лапки, которыми он научился чистить усики после ампутации передних лапок (по Д. Люко, А. Аранда):
I — до ампутации, II —через 12 дней после ампутации, точки под записями — момент прихода импульсов к ганглию, двухфазные потенциалы зарегистрированы от эфферентных волокон данного ганглия
Давно высказанное предположение, что памятный след закрепляется «прорастанием» дополнительных нервных окончаний (Р. Кахал, 1911), позже получило новые подтверждения. Оказалось, что так называемые шипики, несущие синапсы на дендритах корковых клеток, появляются именно в возрасте, когда развивается условно-рефлекторная деятельность. Дополнительные указания на роль шипиков в мехаизмзах памяти дали эксперименты, показавшие интенсивное развитие их синаптического аппарата у животных в результате выработки условных рефлексов. К такому пониманию природы долгосрочной памяти приводит и концепция участия генетического аппарата в пластическом обеспечении функций нервных клеток, согласно которой их интенсивная деятельность при обучении вызывает усиление синтеза белка и процессов роста, в частности, образующих новые синаптические связи. Наконец, зависимость количества синапсов (по показателям электронной микроскопии) от интенсивности функционирования нервных механизмов продемонстрирована в экспериментах с повышением активности синапсов путем предъявления сложных задач или ее снижения путем сенсорной депривации.
По другим предположениям долгосрочная память реализуется облегчением передачи импульсов по существующим синапсам. Это может быть достигнуто стойкими изменениями структуры и свойств синапсов (табл. 3).
Эффективность передачи возбуждения в синапсах в значительной мере зависит от их функционального состояния, определяемого уровнем деятельности. Показано, что усиленное использование синапсов повышает их проходимость для импульсных потоков. При объяснении этого явления учитывается, что ничтожные просветы синаптических щелей соизмеримы с размерами макромолекул мембран, образующих их стенки. Поэтому многократная конвергенция импульсов условного и безусловного раздражителей на синапсах центрального нейрона, вызывая физико-химические изменения его мембран, может, в конечном итоге, сделать синаптическую щель более преодолеваемой.
Физико-химические изменения мембран, возникающие при стойком сдвиге их поляризации, могут служить основой для формирования памятных следов. Такие сдвиги поляризации вопроизведены в экспериментах с созданием зон повышенной возбудимости в коре при помощи постоянного тока и, видимо, имеют место в естественных условиях возникновения доминантных очагов. Опытным путем показано, что создаваемое в доминантном очаге коры состояние возбудимости подпорогового уровня под действием приходящего афферентного возбуждения может достичь порога проявления ответной реакции. Образуется суммационный рефлекс, который при закреплении следов проторения новых путей становится настоящим условным рефлексом. Характерная особенность такого рефлекса состоит в том, что по показателям нейронной активности он протекает в две фазы: торможения и последующей отдачи. Образование таких временных связей происходит преимущественно в нейронах нижних слоев коры.
Одним из механизмов повышения эффективности синапсов могут быть процессы, проявляющиеся в так называемой посттетанической потенциации (Д. Ллойд, 1949). После раздражения афферентных путей синаптическая проводимость мотонейронов спинного мозга остается повышенной в течение 7 мин. Раздражением гиппокампа удавалось продлить такое состояние до нескольких часов. Показано, что гиппокампальная посттетаническая потенциация сопровождается увеличением фокальных потенциалов и укорочением их латентного периода, особенно выраженных, если тетанизации подвергались структуры со свойствами самостимуляции.
Таблица 3. Возможные механизмы устойчивого повышения проводимости в синапсах (по С. Барондес в модификации И.П. Ашмарина)
1. Область изменений в синапсе
2. Состояние синаптической щели
3. Характер изменений
4. Молекулярный механизм
1. Пресинаптическая мембрана
2. Сужение
3. Разрастание нервных окончаний
4. Усиленный синтез белков при запоминании, в дальнейшем их обновление
1. —
2. Без изменения
3. Усиление синтеза и выхода медиатора
4. Дерегрессия части генов медиатора, повышение активности систем трансляции и РНК медиатора
Усиление синтеза медиатора и облегчение выхода из везикул
1. Постсинаптическая мембрана
2. Без изменений
3. Увеличение числа и чувствительности рецепторов
4. Усиленный синтез белков и липидов при запоминании, в дальнейшем их обновление
1. —
2. —
3. Повышение проницаемости и уменьшение скорости расщепления медиатора
4. Измерение соотношений скоростей синтеза компонентов или образование новых компонентов рецептора при запоминании, в дальнейшем поддержание новых отношений. Репрессия генов энзима, расщепляющего медиатор, дерепрессия генов ингибитора этого энзима
1. Локализация не установлена
2. Не известно
3. Повышение проводимости синапса за счет включения неизвестного вещества
4. Усиленный синтез веществ типа антител или других факторов межклеточного «узнавания» (например, полипептиды — коннекторы), при запоминании, в дальнейшем их обновление
Усиленный синтез некоторых антигенов компонента синапса, вызывающих образование соответствующих антител при запоминании, в дальнейшем их обновление
Анализ этих явлений нейронной активности при выработке локальной условной реакции вздрагивания приводит к выводу о том, что для синаптического проторения путей временной связи необходима кроме конвергенции сигнального и безусловного стимулов активация системы, вызывающей модификацию синапсов, что зависит от биологической значимости образования условной реакции. На изолированном нейроне виноградной улитки получены условные изменения свойств его электровозбудимой мембраны путем сочетания стимулов, приложенных к двум ее локусам, в результате чего между ними возникала временная связь.
При изучении возможных причин стойкого повышения синаптической передачи было обращено внимание на медиаторы возбуждения и прежде всего на ацетилхолин. Найдено, что обучение сопровождается повышением активности фермента холинэстеразы, разрушающей ацетилхолин, а вещества, подавляющие действие холинэстеразы, вызывают нарушения памяти. В опытах с обучением крыс доставать пищу из узкой трубки определенной передней конечностью активность холинэстеразы возрастала только в контралатеральном полушарии, что подтверждает ее значение в формировании памятных следов.
Сопоставление данных о взаимосвязанной динамике ацетилхолина и холинэстеразы привело к заключению, что повышенная проводимость может возникать и устойчиво поддерживаться некоторым равновесным отношением их синтеза. Избыточная активность холинэстеразы без возрастания количества ацетилхолина не позволяет ему достичь концентрации, необходимой для деполяризации постсинаптической мембраны синапса, а увеличение количества ацетилхолина при малой активности холинэстеразы приводит к глубокой неколеблющейся деполяризации — в обоих случаях проведение импульсов через синапс оказывается блокированным. Существует предположение, что ацетилхолин может транспортироваться через синаптическую щель по транссинаптическим тяжам, которые образуются из соединения белковых мицелл, формирующихся на поверхности пре- и постсинаптической мембран.