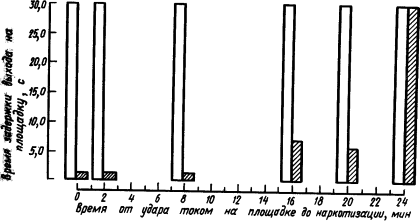
Рис. 20. Влияние наркоза на закрепление у мышей рефлексов избегания выхода на площадку, находящуюся под напряжением (по И. Абт и др.):
белыми столбиками изображено проявление рефлекса до наркотизации, заштрихованными — после наркотизации в разные сроки; остальные объяснения см. в тексте
На основании опытов с обезьянами было высказано предположение, что в условно-рефлекторной деятельности проявляются оба вида памяти: при отсроченных условных рефлексах — преимущественно краткосрочная, при следовых условных рефлексах — долгосрочная. Поэтому более вероятно, что механизмы краткосрочной и долгосрочной памяти тесно связаны между собой и являются последовательными этапами единого процесса, где памятный след, менее устойчивый на первом этапе, на втором этапе становится прочным. О природе двух видов памяти высказывают разные предположения, но, несмотря на интенсивные исследования, единой общепризнанной теории памяти пока не существует.
Возможные механизмы краткосрочной памяти. Наиболее распространено предположение, что краткосрочная память реализуется в виде многократной циркуляции потоков импульсов по замкнутым цепям нервных клеток. Основанием для такой гипотезы реверберации возбуждения были морфологические данные о наличии в сетях нейронов множественных соединений, образующих по петлям обратных связей круговые пути (Р. Лоренте де Но, 1934). Сочетания условного и безусловного раздражений активируют эти пути и многократное возбуждение нервных клеток, по которым протекают потоки импульсов, постепенно вызывает в них стойкие изменения, образующие долгосрочную память. Действительно, все воздействия, нарушающие краткосрочную память, прерывают возникающую циркуляцию потоков импульсов по замкнутым нервным путям.
Однако дальнейшие исследования показали, что в круговых путях нервных сетей находятся не только возбуждающие, но и тормозящие клетки, которые препятствуют «зацикливанию» циркуляции импульсов. На рис. 21 показана такая схема сети, в которой клетка 1 с клеткой 1А могла бы образовать круг реверберации импульсов, обусловливающей ритмическое возбуждение клетки Б. Но достижение порога возбуждения тормозящей клетки 1Б разрывает этот круг, затормозив клетку 1А.
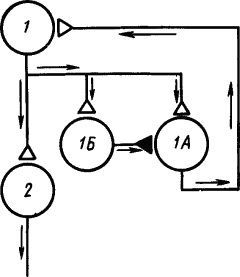
Рис. 21. Нервная цепь с замкнутыми нейронными цепями, по которым могут циркулировать потоки импульсов (по Э. Кенделу):
1 — корковый нейрон, получающий афферентные сигналы, 1А и 1Б — промежуточные нейроны, 2 — эфферентный нейрон; белыми треугольниками изображены возбуждающие синапсы, черными — тормозящий, стрелками показано направление движения потоков импульсов
Краткосрочная память не требует для своей реализации синтеза информационных (матричных) молекул РНК и ДНК. Она и не могла бы обеспечиваться этими биохимическими процессами как генетическая память, поскольку включение в полимер ДНК или РНК нуклеотидов происходит в сотни и тысячи раз медленнее, чем это необходимо для запечатления в них полной информации о текущих событиях. Вместе с тем при любом понимании ее природы, в том числе и реверберационном, речь идет о протекании потоков импульсов по новым для них путям, т.е. о временной синаптической проводимости.
Для объяснения механизма синаптической проводимости была предложена гипотеза о взаимодействии медиатора ацетилхолина с рецепторными белками постсинаптической мембраны и Na+, K+-АТФ-азой при разных концентрациях ионов Na и Ca. При интенсивной импульсации выброшенный из везикул пресинаптической мембраны ацетилхолин связывается с рецепторными белками постсинаптической мембраны, подавляет деятельность ионных насосов, деполяризует мембрану и выключает Na+, K+-АТФ-азу. Однако возникающие при этом потенциалы возбуждения и уменьшение отношения Na+ и K+ приводят к высвобождению ацетилхолина и последующему разрушению его холинэстеразой, что может восстанавливать активность Na+, К+-АТФ-азы на более высоком уровне.
Временное повышение синаптической проводимости связывают также с высвобождением Ca2+ под действием нервных импульсов в пресинаптических окончаниях, который вызывает усиленное сокращение везикул и выбрасывание медиатора, или с повышенной чувствительностью белков деполяризованной мембраны к протеолитическим ферментам, делающим ее более проходимой для медиатора. В пользу последнего предположения свидетельствует возрастание активности протеаз в ткани мозга животных при выработке условного рефлекса.
Исходя из результатов электрофизиологических исследований было высказано мнение, что краткосрочная память образуется за счет посттетанической потенциации, поддерживаемой реверберующими потоками импульсов. Посттетанической потенциацией, или посттетаническим облегчением, называют увеличение числа проходимых синапсов после ритмического раздражения пресинаптических волокон. На то, что это явление зависит от изменений в их окончаниях, а не в постсинаптических структурах, указывает отсутствие такого эффекта при антидромной стимуляции (рис. 22). Из рис. 22 видно, что в данном случае облегчение длится около двух минут и что антидромная стимуляция оказывается безрезультатной.
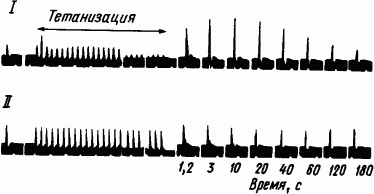
Рис. 22. Посттетаническое временно́е облегчение проведения через звездчатый ганглий кошки (по М. Ларраби, Д. Бронк):
I — ортодромная стимуляция, II — антидромная стимуляция, объяснение см. в тексте
Несмотря на различия предположений о природе краткосрочной памяти, их объединяет рассмотрение механизма памяти как возникновения непродолжительных обратимых физико-химических свойств мембран и динамики медиаторов в синапсах, временно перестраивающих поведение нервных сетей. Эти изменения неизбежно связаны со сложной цепочкой далеко идущих, сравнительно более стойких перестроек метаболизма нервной клетки. Поэтому кратковременная память может оставлять более длительный скрытый след, основе которого в некоторых случаях она способна восстанавливаться после действия амнестического агента. Углубление и структурно-метаболическое закрепление этих следов, по-видимому, является механизмом перехода краткосрочной памяти в долгосрочную.
В отличие от краткосрочной долгосрочная память хранит следы пережитых событий неопределенно долго. Собственно она и является носителем жизненного опыта животного; знаний, приобретенных человеком. О субстрате и способе закрепления долгосрочных памятных следов существуют разные мнения.
Долгосрочная память как возникновение новых межклеточных связей и улучшение проведения по уже существующим связям. Не вызывает сомнений, что в основе всякого обучения лежит процесс проторения путей возбуждения. Наглядной иллюстрацией того, что в результате обучения потоки нервных импульсов легче и быстрее преодолевают синаптическую задержку, может служить модельный опыт с тараканом: после ампутации передних лапок, которыми он чистит усики, таракан обучается пользоваться для этой цели задними лапками. При этом время синаптической задержки моторных импульсов к задним лапкам резко уменьшается (рис. 23).
Исследование динамики активности нейронов виноградной улитки при выработке оборонительного условного рефлекса показало, что повышение эффективности синапсов происходит на входах командных нейронов, реализующих защитные движения.